Association Martiniquaise des Amis des Volcans Verts de la Caraïbe

La Martinique appartient au point chaud de biodiversité des Caraïbes.
Un point chaud ou hot-spot de biodiversité est une zone géographique contenant au moins 1500 espèces végétales endémiques mais qui a déjà perdu au moins 70 % des espèces présentes dans leur état originel.
La surface totale des points chauds ne représente que 2,3 % de la surface de la Terre. A lheure actuelle, 34 points chauds ont été répertoriés. Plus de 50 % des espèces végétales et 42 % des espèces de vertébrés terrestres vivent dans ces points chauds.
Les îles des Caraïbes, avec leurs deux aires biogéographiques (Grandes Antilles et Petites Antilles), correspondent à l'un de ces points chauds.
Au sein de l'aire biogéographique des Petites Antilles, la Martinique occupe une place prépondérante, elle est l'île qui présente la plus grande biodiversité en espèces arborées indigènes. Elle est également l'île qui présente le plus grand nombre d'espèces arborées endémiques d'une île. Pour les autres espèces arborées endémiques des Petites Antilles, elle partage cet endémisme avec une ou plusieurs îles des Petites Antilles. Elle est donc au cur de la biodiversité des Petites Antilles. et est talonnée par ses voisines : Sainte-Lucie, La Guadeloupe et La Dominique.
Des hypothèses ont été avancées pour tenter d'expliquer la suprématie des îles du centre de l'archipel, il nous appartient de faire la part des choses en prenant en compte les nouvelles données relatives à l'approche biogéographique de la Caraïbe.
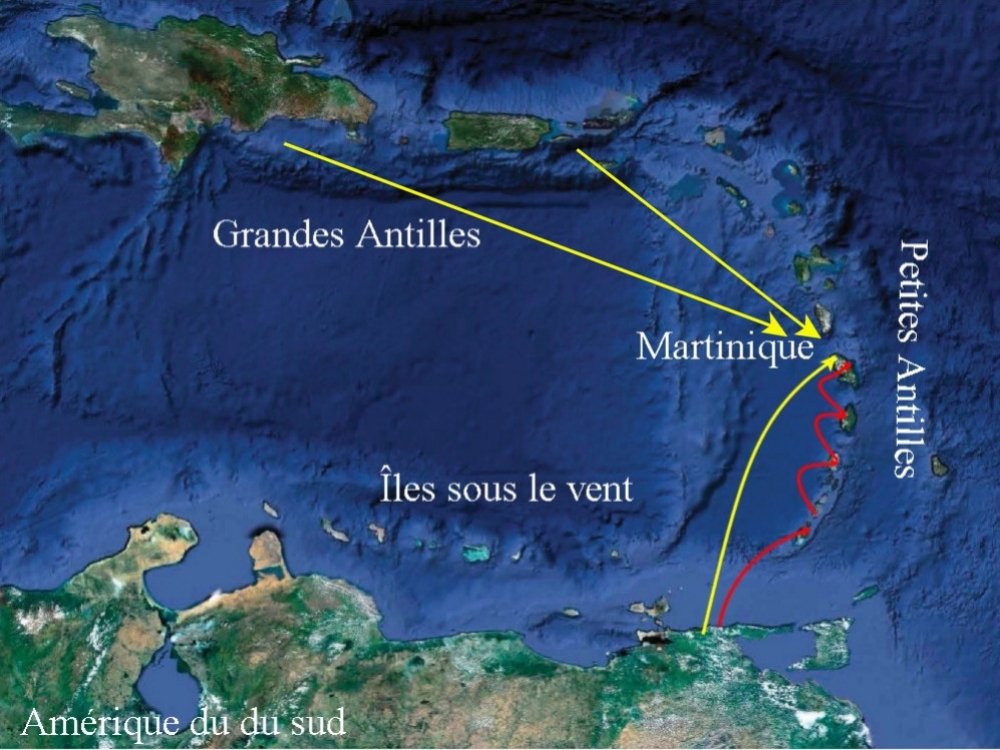
Dans un passé récent, il était admis que : "Larc des Petites Antilles navait jamais été relié au continent
américain ni aux Grandes Antilles, géologiquement plus anciennes et que les
îles elles-mêmes ou leurs ébauches ancestrales avaient émergé isolément
dans lOcéan Atlantique et étaient depuis longtemps séparées les unes des
autres". Pour expliquer la dispersion dîle en île, deux types de migration sur "bois flottés" avaient alors été envisagées, la migration par "saut de puce" dune île à lautre et la migration par "saut de grenouille" qui permettait dexpliquer des répartitions atypiques. Qu'en est-il aujourd'hui ?
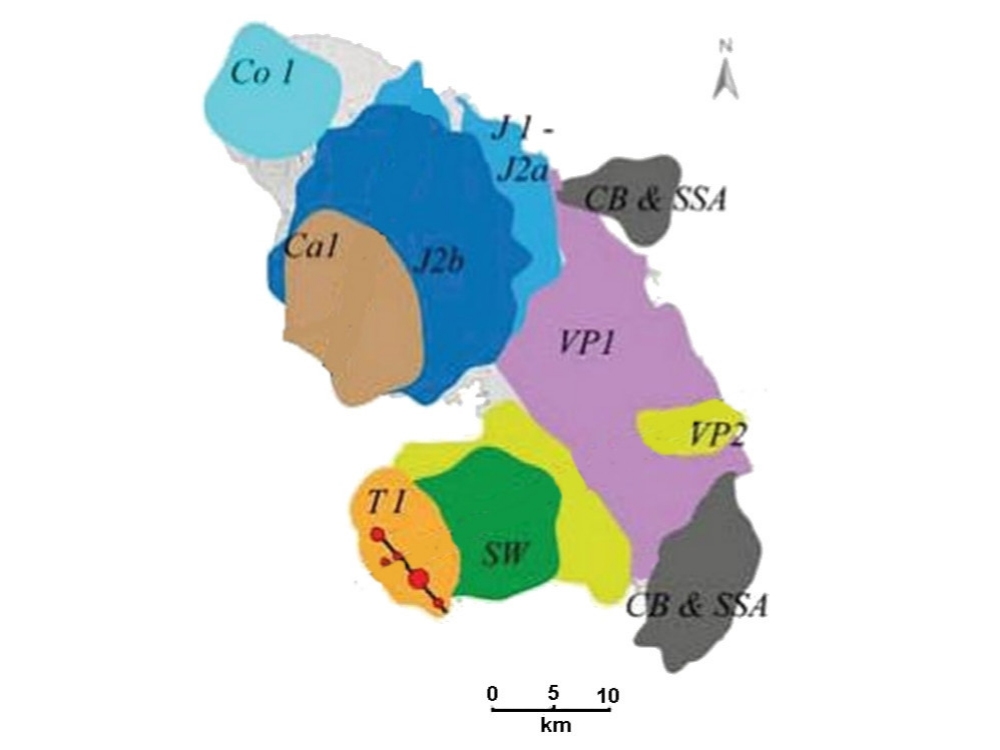
La Martinique est une île archipel : elle est constituée de plusieurs
petites îles volcaniques qui se sont formées les unes à côté des autres
avant d'être reliées par des ponts volcaniques.
Elle est la seule île des Petites Antilles a présenter trois arcs volcaniques juxtaposés, avec un déplacement régulier de l'activité volcanique de l'est vers l'ouest depuis 25 Ma
Elle est très probablement l'une des plus vieilles îles volcaniques actives du monde.
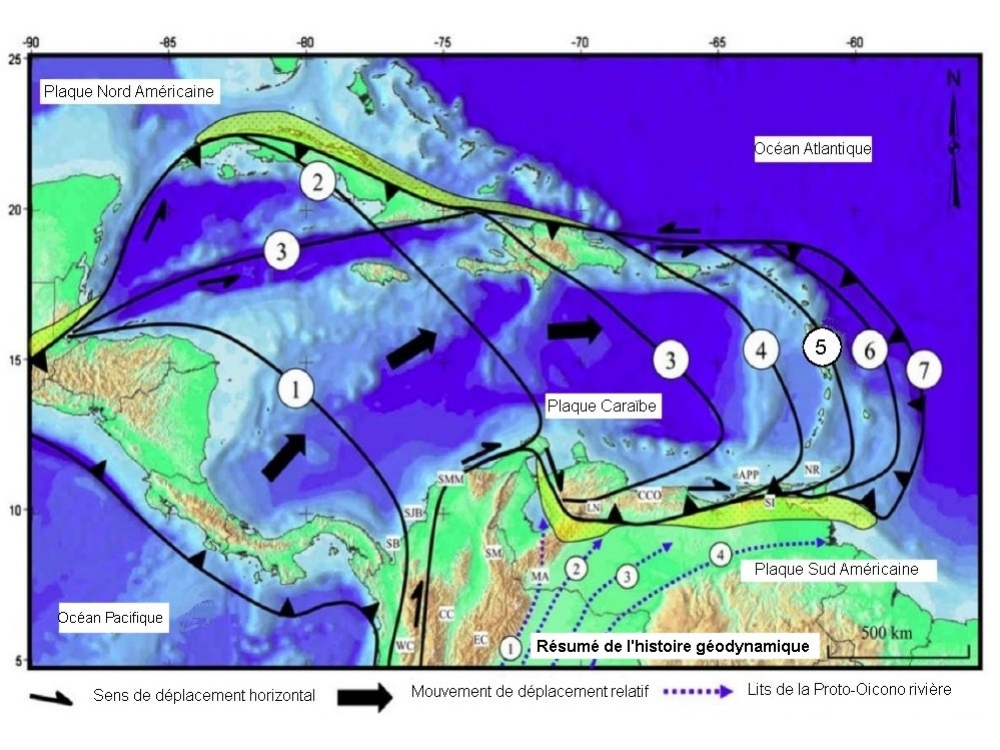
Le soubassement de la Martinique est constitué d'un fragment de la Ride dAvés qui a pris naissance il y a 120 Ma dans l'Océan Pacifique.
Cette ride a fait partie d'un arc volcanique actif appelé l'Arc Mésozoïque Caraïbe qui s'est régulièrement déplacé de l'ouest vers l'est avec le coulissage de la Plaque Caraïbe entre les deux Amériques.
Après l'ouverture du Bassin de Grenade, l'Arc ancien des Petites Antilles prend naissance sur ce soubassement volcanique d'âge mésozoïque.
Au sein du hot spot de biodiversité des Petites Antilles, la Martinique apparait comme étant lîle présentant la plus grande biodiversité en espèces arborées autochtones.
Elle est la plus riche en espèces arborées endémiques dune île, la plus riche en espèces arborées endémiques partagées avec les autres îles des Petites Antilles, la plus riche en espèces arborées endémiques partagées avec les Grandes Antilles.
Elle est donc au cur de la biodiversité des Petites Antilles.
Une telle richesse ne peut être le fruit du « hasard de sauts de grenouille », s'il en avait été ainsi, la Martinique se serait comportée par le passé comme ayant été un véritable aimant d'attraction de biodiversité.
Recherchons des indices qui pourraient nous permettre davancer des propositions explicatives plus rationnelles.

Cette étude montre que la Presquîle du Sud-ouest est au cur de la biodiversité en espèces arborées endémiques des étages xéro-mésophiles des Petites Antilles. La quasi totalité de ces espèces ont une origine sud-américaine, pour expliquer cette singularité, il faudra rechercher d'autres voies de migration que celles qui existent aujourd'hui. Elle n'a pas pu se faire en passant par les îles situées au sud (Saint Vincent et Grenade) qui sont beaucoup plus pauvres.

Cette étude montre que, bien quelle soit plus éloignée des Grandes Antilles que les grandes îles du nord (Guadeloupe, Dominique) la Presquîle du Sud-ouest a eu par le passé des échanges privilégiés en direct avec les Grandes Antilles.
Ces échanges ont du se faire par intermédiaire de l'Arc externe avant l'immersion des îles du nord qui a effacé toute mémoire vivante de cette connexion passée.

Des ossements et des dents d'un "rat géant" Amblyrhiza inundata ont été trouvés dans des formations sédimentaires des îles de Saint-Barthélemy, Saint Martin et d'Anguilla.
La présence de ces fossiles sur les îles du nord de l'arc ancien
permet d'émettre l'hypothèse d'une connexion territoriale entre les
Grandes Antilles, l'arc ancien externe des Petites Antilles, la Ride
d'Avés et l'Amérique du sud.

Il est possible d'établir les liens de parenté entre différentes espèces animales et végétales en établissant le "code bar" de leur ADN. Ces études menées sur les mabouyas des litières de Martinique, les châtaigniers des Petites Antilles et la grenouille dendrobate de la Montagne Pelée laissent à penser que l'archipel miocène de Martinique a joué un rôle déterminant dans l'apparition de nouvelles espèces endémiques de Martinique et des Petites Antilles.
L'ensemble de ces études permettent de proposer au moins trois voies de migration pour expliquer la colonisation de la Martinique à partir de l'Amérique du sud.
Ces voies se sont relayées au cours du temps pour déboucher sur des populations animales et végétales dont la composition exprime la synthèse de cette histoire géologique.
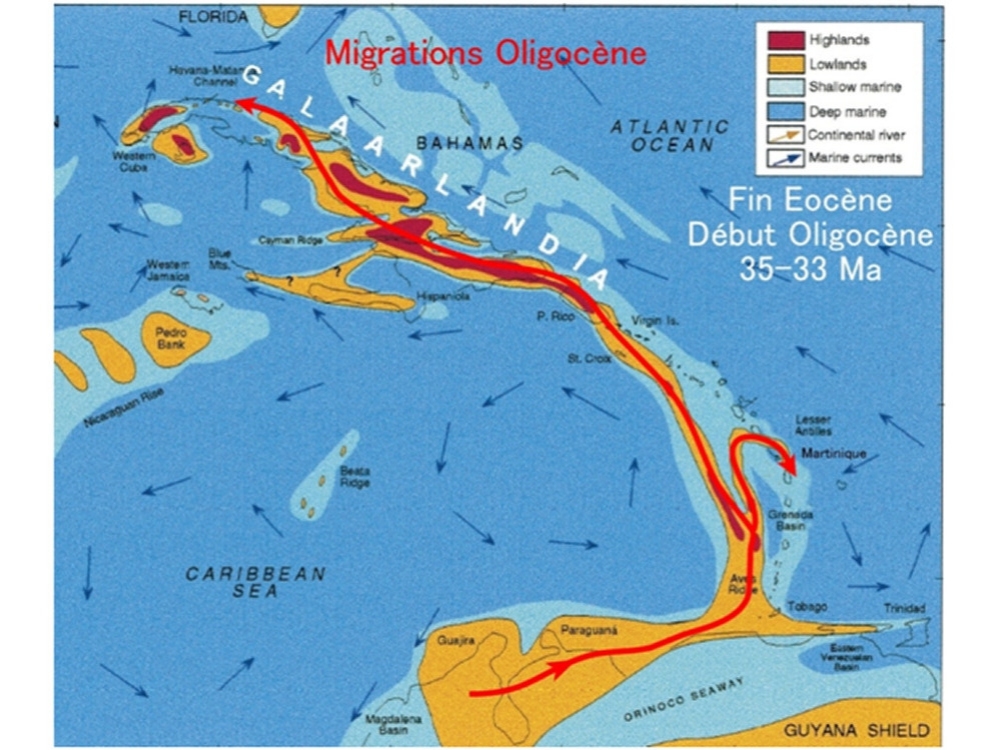
Au début de l'Oligocène (35 à 33 Ma) la glaciation de lantarctique conjuguée aux effets de la tectonique compressive a provoqué lémersion de la Ride dAvés qui a alors formé un pont terrestre reliant les Antilles à lAmérique du sud.
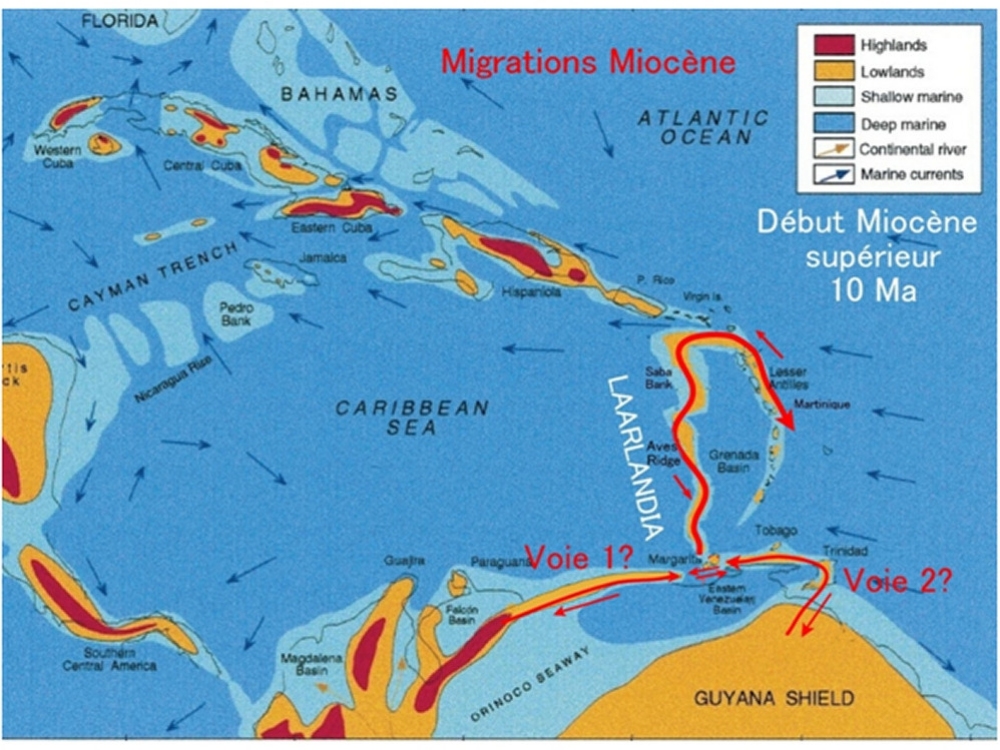
Une nouvelle glaciation conjuguée aux nouvelles contraintes tectoniques
du Miocène moyen a pu provoquer une nouvelle émersion totale ou
partielle de la Ride dAvés désormais séparée des Grandes Antilles par
le Passage dAnegada.
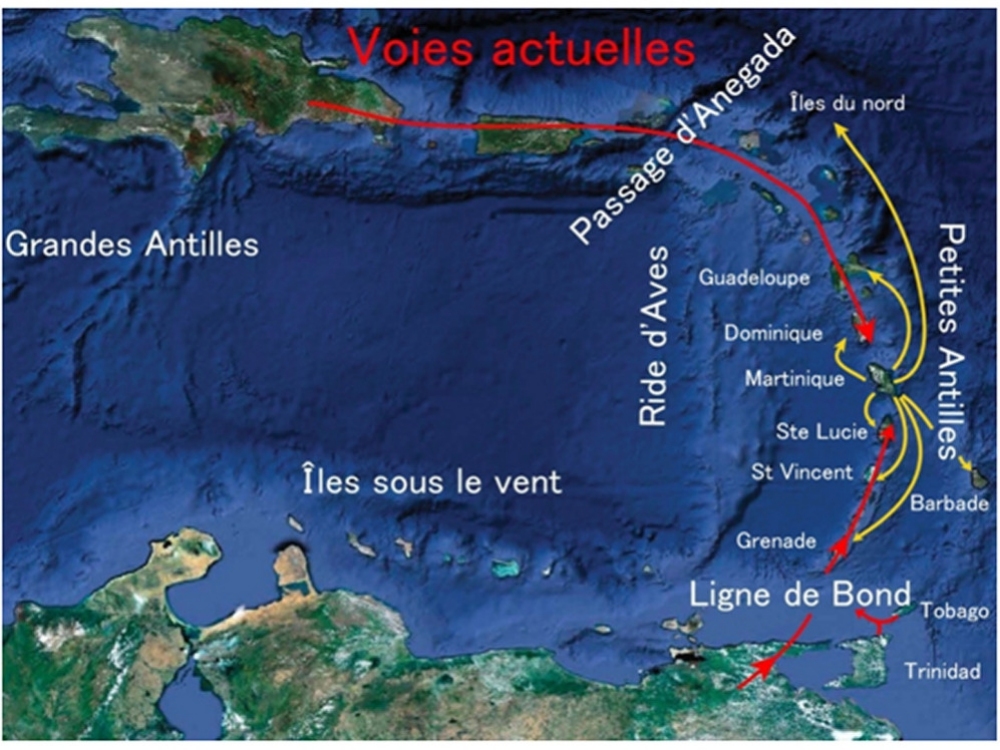
Ces migrations se font d'île en île par saut de puce. Elles ont pu être véhiculées par les cyclones et les bois flottés. Les nombreuses déstabilisations de flanc qui ont émaillées l'histoire des volcans des Petites Antilles ont très largement contribué à ces échanges.
La Martinique, île archipel, résume bien les grandes étapes de lévolution géologique de lest caraïbe.
Elle a été construite par le fonctionnement de plusieurs arcs volcaniques qui se sont déplacés et relayés au cours du temps. Elle est la seule île de la région à exposer à sa surface de manière éclatée une histoire vieille de plus de 25 Ma sur un soubassement volcanique vieux de 120 Ma.
Au cours de cette longue histoire de l'est Caraïbe, les îles volcaniques formées au niveau des arcs nont eu de cesse de jouer au yoyo, avec des phases démersion suivies de phases dimmersion effaçant toute trace de biodiversité terrestre passée.
Ces soulèvements ont été si importants que ces îles ont parfois formé de véritables ponts reliés au continent sud américain amenant ainsi des vagues dessaimage de la biodiversité sud-américaine.
Tout au long de cette histoire, certaines de ces îles ont toujours gardé la tête hors deau. Lisolement géographique ainsi réalisé a été le principal moteur dune spéciation conduisant à un endémisme exceptionnel. Ces îles se répartissent en deux groupes, les Grandes Antilles au nord et les petites îles de lArche de Noé au sud.
Létude de la biodiversité actuelle laisse donc à penser que le territoire correspondant à ces îles de lArche de Noé anciennement connectées à lAmérique du sud, se limite à lArchipel Miocène de la Martinique et à Sainte-Lucie. Contrairement à ce qui sest passé pour les Grandes Antilles définitivement isolées depuis la fin de lOligocène (33 Ma), cet archipel a de nouveau été réalimenté par des vagues successives de biodiversité venant directement dAmérique du sud.
Compte tenu de la faible représentation de cette biodiversité dans les
autres îles du sud, on peut penser que leur territoire n'ont pas été connectés
au continent sud américain. La différence entre la Martinique et Sainte-Lucie trouverait alors son explication dans le fonctionnement de l'Arc intermédiaire. A Sainte-Lucie, la superposition des formations volcaniques aurait exercé un effet destructeur sur la biodiversité initiale et limité la spéciation par isolement géographique (absence d'archipel).
Au niveau de l'Archipel Miocène de Martinique, cette biodiversité exceptionnelle sest trouvée concentrée sur un très petit territoire constitué de petites îles très proches les unes des autres. Isolement géographique, dispersion et spéciation ont renforcé le caractère explosif de cette biodiversité primaire et aboutissent ainsi sur des ratios biodiversité/surface tout à fait exceptionnels.
La Presquîle du Sud-ouest, figure emblématique de cet Archipel Miocène, véritable concentré de cette biodiversité primaire a su garder encore aujourd'hui - pour combien de temps encore - des témoins vivants de cette histoire exceptionnelle.
A ce titre, elle doit absolument être protégée et valorisée.